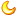|
精子的"前世今生" 精子是有性生殖、异性配偶形式生殖方式中的雄性生殖细胞(或配子),一般情况下, "女性"生殖细胞较大,而"男性"生殖细胞较小。动物产生的精子可"游动",其尾巴被称为鞭毛,,而一些红藻和真菌则产生"非活动性"的精子细胞。开花植物的花粉中含有不"活动"的精子,而蕨类和裸子植物等一些初等植物则具有"活动"的精子。

精子细胞的形成在羊膜动物(爬行动物和哺乳动物)中发生在睾丸的生精小管中,该过程涉及几个连续的精子细胞前体的产生。首先是精原细胞,然后分化为精细胞。然后,精母细胞进行减数分裂,使其染色体数目减少一半,从而产生精子。之后,精子成熟,精子的尾部形成一条尾巴或鞭毛,从而形成成熟的能动精子细胞。成熟精子形成的整个过程从开始到结束大约需要3个月的时间。 
精子细胞不能分裂并且寿命有限,但是在受精过程中与卵细胞融合后,新的生命体便开始发育。人的精子细胞是单倍体,它的23条染色体可以与雌卵的23条染色体结合,形成二倍体细胞。在哺乳动物中,精子被储存在附睾中,并在射精过程中以精液的形式从阴茎中释放。正常成年男性一次射精量大约在2-5ml左右,一次射出的精子数目可达3-5亿。。

精子发生过程基因表达的复杂性在哺乳动物所有器官中,睾丸表达的基因数目最多。这一生物现象也长期困扰着分子生物学家。最近一项有关人类、小鼠精子发生的单细胞转录组学研究表明,精子细胞基因的广泛表达有助细胞检查并修复基因组DNA损伤,从而维持雄性种系DNA序列的完整性。 研究人员还发现,与精子发生过程中相关基因突变率相对低,并且基因序列在种群中具有较低的多样性。与精子发生过程无关的基因在时间尺度上进化更快。总的来说,"转录扫描"(transcriptional scanning)以作用于特异性基因的方式调节种系突变率,维持大部分基因的DNA序列完整性,但允许部分特定基因中快速变异、进化(如免疫相关基因)。 
多年以来,睾丸转录组一直被认为是最复杂的。据有关研究报道,睾丸中广泛发生的基因转录,覆盖了人类及其他物种超过80%的蛋白质编码基因。科学家相继提出一些假设来解释这一观察结果。

精子基因高表达的理论学说第一个假说观点认为:基因广泛表达可能是为了弥补部分基因表达产物异常。 但是,更复杂的器官(例如大脑)细胞类型虽然明显更多,却没有显示相应基因数量大量表达。 此外,最近有研究表明,雄性小鼠的生育能力,并不需要许多睾丸组织特异基因以及进化上相对保守的基因发挥功能而维持。睾丸中的转录组和蛋白质组之间显著的不一致进一步支持了这一观点,即广泛的基因转录并不一定会通过"中心法则"导致功能性蛋白质合成。

第二个假说观点认为:在精子发生的整个过程中,大量染色质重塑过程中出现的"转录失误"(leaky transcription)。 该模型预测在精子发生的后期阶段(当基因组的染色质变化最大时)会有更多的表达,但是这与先前的观察结果矛盾。 另外,鉴于广泛的基因转录对能量的高需求,"转录失误"(leaky transcription)理应受到更严格的限制,缩减转录基因的数目似乎才更加符合生命维持逻辑。

科学家提出的第三种模型:"转录扫描"(transcriptional scanning)模型。这一理论认为,睾丸内基因的广泛转录可调节基因的进化速度。使用人和小鼠睾丸细胞进行scRNA-Seq分析,证实了广泛的基因转录确实来自生殖细胞,而不是来自体细胞和生殖细胞两部分。 研究发现,与未表达的基因相比,精子发生表达的基因在人群中的遗传变异(germline variants)更少,并且这些基因上的转录偶联修复(TCR)标志(signature)可以来解释观察到的有偏差的生殖系变异模式。该模型认为,在精子发生过程中,广泛的基因转录可视为一种DNA扫描机制(transcriptional scanning)。 
在"扫描"过程中通过转录偶联修复(TCR)系统地检测和修复庞大的基因组中存在的DNA损伤,从而降低种系突变率,并最终降低基因进化的速率,使亲子代之间保持一定的遗传稳定性。在雄性种系中未表达的基因并不是随机不确定的。事实情况是,这些未表达基因主要为感觉和免疫/防御系统相关基因,这与先前研究观察到这类基因进化得更快的结论相一致。

但是,在高表达的基因子集中,转录偶联损伤(transcription-coupled damage ,TCD)远远超过了转录偶联修复(transcription-coupled repair ,TCR)的作用,这些基因富含与精子发生有关的功能,这也暗示了转录偶联损伤(transcription-coupled damage ,TCD)在种系突变率调节中的作用。总的来说,"转录扫描"( transcriptional scanning)模型揭示了迄今为止在DNA修复中尚未得到重视的方面,这一理论模型更加侧重于整个基因组的基因进化速率。
| 







 雷达卡
雷达卡 发表于 2020-4-20 10:26:50
发表于 2020-4-20 10:26:50
 提升卡
提升卡 置顶卡
置顶卡 沉默卡
沉默卡 喧嚣卡
喧嚣卡 变色卡
变色卡 千斤顶
千斤顶 显身卡
显身卡

 发表于 2020-4-20 10:29:19
发表于 2020-4-20 10:29:19